
Si sono osservati aumenti delle concentrazioni plasmatiche di treonina a fronte di riduzioni della proteina grezza nella dieta del pollo, ma l’ampiezza di queste variazioni pare correlata a peggiori conversioni. In questo studio si cerca di spiegare il meccanismo che porta a questi alti livelli plasmatici.
Introduzione
La treonina è l’amminoacido (AA) scoperto più di recente, ma al contempo è anche il terzo AA limitante nella dieta del pollo. C’è un notevole interesse nel ridurre il contenuto di proteina grezza (CP) nel mangime del pollo, perché darebbe diversi vantaggi, tra cui una minore dipendenza dalle importazioni di soia. Negli anni ’80 si vide che le concentrazioni di treonina libera nel plasma sistemico variavano fino all’87% (1635 rispetto a 876 nmol/L) nelle femmine di 42 giorni in seguito a una riduzione della CP da 183 a 159 g/kg. Inoltre si sono visti comportamenti simili della treonina fino al 116% (1093 rispetto a 505 μmol/L) a livello plasmatico, dopo una riduzione della proteina grezza (da 200 a 156 g/kg di CP) in 4 livelli dietetici di CP in questa prova. Pertanto ci si interroga circa la genesi e la rilevanza di questi picchi di treonina a livello plasmatico a seguito di diete ipoproteiche.
Discussione
La treonina è un amminoacido essenziale, quindi esiste la possibilità che i suoi livelli plasmatici elevati siano dovuti a un’alterazione della funzionalità degli enzimi epatici, in grado di catabolizzarla. La treonina viene convertita ad acetaldeide e glicina dalla treonina aldolasi (TA), in un α-chetobutirrato dalla treonina deidratasi (TH) e in acetil-CoA e glicina dalla treonina-3-deidrogenasi (TDH). Inoltre si è visto che l’attività epatica della TDH (88%) è dominante nelle specie avicole, mentre la TH (93%) lo è nei ratti. Visto che TDH metabolizza la treonina in acetil-CoA nello studio si è concluso che la treonina è un AA chetogenetico negli avicoli, a differenza dei mammiferi, dove viene considerato esclusivamente glucogenetico. Ne segue che la genesi di livelli elevati di treonina libera nel plasma derivi dall’ipoattività della TDH epatica. Teoricamente, la treonina è un precursore della glicina, ma non sarebbe questo il caso se l’attività della TDH venisse diminuita. Alcuni studi mostrano che aumentare la concentrazione di treonina plasmatica è associato in maniera lineare (r=-0,608; P<0,001) a un calo della concentrazione di glicina, il che indica che la treonina non viene catalizzata in glicina.
Diversi studi hanno valutato le conseguenze di vari livelli dietetici di proteine, amminoacidi e treonina sull’attività della TDH, ma senza portare a conclusioni definite. Sembrerebbe infatti che l’attività della TDH epatica venga influenzata dai livelli di proteine nella dieta o da altri amminoacidi, piuttosto che dalla treonina medesima. La treonina è l’AA dominante della mucina aviare ed è pure presente in altre secrezioni endogene intestinali. Di conseguenza, circa il 30% della treonina viene destinata alla mucosa intestinale e non entra nel circolo portale. Inoltre, viene anche utilizzata per la formazione della livrea e per mantenere la immunocompetenza, oltre che essere incorporata nelle proteine. Ciononostante restano oscure le ragioni per cui una riduzione proteica nel mangime faccia aumentare i livelli di treonina.
Il catabolismo della treonina da parte dell’enzima TDH genera acetil-coenzima A, che è un attore del metabolismo centrale, capace di influenzare l’attività di enzimi multipli. È stata anche studiata l’inibizione dell’attività epatica della TDH nei ratti, con un’attenzione agli acidi grassi, ma si è concluso che la TDH sia il target di una inibizione selettiva a ritroso (feed back) da parte di tutti i composti derivanti dall’acetil CoA, come prodotto finale. Comunque , il glucosio può venire metabolizzato per generare acetil CoA , il che può essere fondamentale. Le diete a ridotta concentrazione proteica tipicamente contengono più granaglie, quindi più amido, e un rapporto alterato amido:proteina. Ridurre i livelli proteici potenzialmente aumenterebbe quelli di glucosio, rispetto ai mangimi convenzionali. Quindi pare che un aumento delle concentrazioni di acetil-CoA derivi da livelli maggiori di glucosio in soggetti alimentati con diete ipoproteiche, il che limita l’attività della TDH e causa quindi questi picchi di concentrazioni plasmatiche di treonina.
Conclusioni
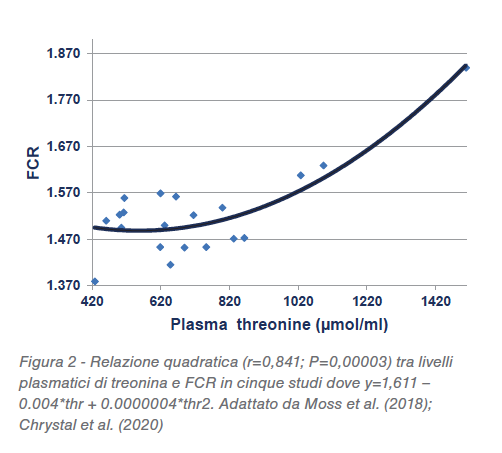
Sono stati formulati 5 livelli dietetici diversi di proteina, da 213,5 a 166,5 g/kg. Esiste una relazione quadratica (r=0,702; P<0,005) tra CP nel mangime e livelli plasmatici di treonina, che comincia ad aumentare quando la proteina grezza cala sotto ai 204,4 g/kg, come mostrato nella Figura 1. Dalla regressione quadratica si può dedurre che una diminuzione da 210 a160 g/kg potrebbe generare un 87,9% di aumento (1090 rispetto a 580 μmol/L) di treonina plasmatica libera. Esiste anche una relazione quadratica (r=0,841; P<0,0001) tra i livelli di treonina plasmatica e il tasso di conversione dove alti livelli di treonina sono associati con livelli di efficienza della conversione più bassi come mostrato in Figura 2. Inoltre c’è una relazione lineare significativa tra le concentrazioni di amido nella dieta (r=0,522; P<0,025) e il rapporto amido:proteina (r=0,623; P<0,005) con livelli plasmatici di treonina.
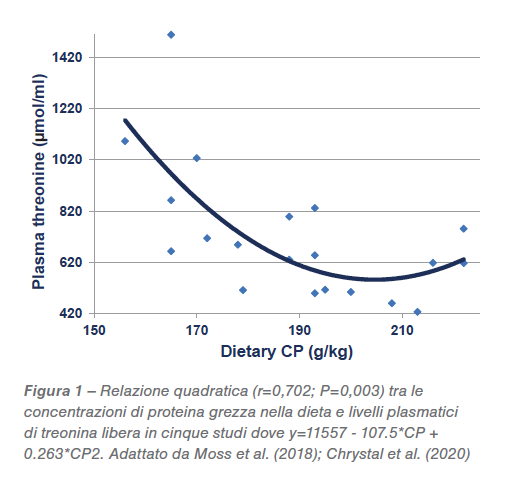
In conclusione, queste relazioni non convalidano il ragionamento, tuttavia sono del tutto coerenti con la premessa che l’acetil-CoA derivato dall’amido e dal glucosio sta limitando l’attività della treonina-3-deidrogenasi e generando elevati livelli plasmatici di treonina. Che i picchi di treonina siano associati al deterioramento del rapporto di conversione FCR non è probabilmente un rapporto di causa-effetto ma può riflettere un disturbo fisiologico sottostante. Sembra che una strategia che potrebbe diminuire le concentrazioni elevate di treonina plasmatica dovrebbe limitare gli aumenti nelle concentrazioni di amido e il rapporto amido:proteina nelle diete ipoproteiche. È stata valutata la possibilità di mantenere costante il rapporto amido:proteina con qualche vantaggio. Una riduzione da 1,68 a 1,41 in diete con 205 g/kg di CP a base frumento, ha aumentato il peso del 10,37% (2161 rispetto a 1958 g/capo), migliorando anche il consumo del 3,1% (3492 rispetto a 3387 grammi/capo) e la conversione del 4,04% (1,6 rispetto a 1,684) a 35 giorni. In questa prova, sostituire il pannello di soia (475 g/kg) con soia non sgrassata (360 g/kg) per ridurre il contenuto di proteina grezza della dieta, riducendo al contempo l’inclusione di frumento da 607 a 502 g/kg, ha portato la concentrazione di amido da 347 a 288 g/kg, mentre la proteina grezza CP è rimasta sostanzialmente invariata (da 206 a 205 g/kg).
Le concentrazioni plasmatiche di treonina libera rappresentano un campo di ricerca davvero interessante. In ratti nutriti con una dieta con AA derivati da caseina, i livelli di treonina aumentavano del 36% (424.4 rispetto a 312.0 μmol/L) dopo 3 ore dal pasto. Ma quando si somministrava una dieta con amminoacidi misti, la concentrazione aumentava fino a 60,5% (760.4 versus 473.7 μmol/L). Si tratta di una differenza del 38% che ci porta a concludere che amminoacidi sintetici nella dieta del pollo possano in qualche modo aumentare i livelli di treonina.
Noi riteniamo che occorra ridurre il rapporto amido:proteina nelle diete ipoproteiche. Ciò potrebbe migliorare le performance e diminuire l’eccesso di grasso di deposito, che si verifica nei polli alimentati con diete a bassa proteina grezza. Date queste premesse, è possibile che i maggiori livelli plasmatici di treonina possano diminuire, contestualmente al miglioramento delle performance.
Bibliografia disponibile su richiesta
Dagli Atti dell’Australian Poultry Science Symposium 2021





