
Il rilevamento delle malattie infettive viene solitamente effettuato o tramite l’isolamento del patogeno o della risposta immunitaria che stimola. Le metodiche diagnostiche sono molte e non esistono metodi standardizzati tra i vari laboratori diagnostici (tranne che per i test NAHLN). I risultati vengono paragonati tra i laboratori con difficoltà, anche per le diverse metodiche, i ceppi o i patogeni usati, in particolare per i test sierologici. Inoltre, con l’introduzione delle metodiche molecolari, che sono notevolmente sensibili, bisogna interrogarsi sui risultati positivi e sul loro significato.
Coltura
Il focus di questo lavoro riguarda il rilevamento dei patogeni e, come abbiamo detto, esistono diversi metodi. Il primo (storicamente considerato quello di riferimento) è la coltura, che è una vera coltura per batteri e funghi e un isolamento per i virus. La cosa importante da considerare, per questa metodica, è la vitalità del microrganismo. Se il campione clinico non è stato gestito correttamente, non rileveremo alcun microrganismo con questa metodica. Inoltre, se cerchiamo di isolare un batterio, è importante far sapere al laboratorio quale microbo si stia cercando. Si usano terreni speciali per determinati microrganismi, come il Mycoplasma. Un buon campionamento è fondamentale per la coltura; il rilevamento di flora ambientale contaminante infatti non favorisce la diagnosi. La cosa interessante della coltura è che si tratta di una metodica piuttosto aspecifica. Occorrono i terreni giusti (per batteri o funghi) oppure le uova /o colture cellulari (per i virus) per trovarli; comunque, non tutti crescono in vitro, oppure alcuni sono difficili da coltivare, quindi altre metodiche sembrano migliori. Poi, l’isolamento virale può richiedere settimane per dare evidenze, e, come per i batteri la sua identificazione richiede ulteriori passaggi, come l’uso di anticorpi specifici, che si legano al virus, oppure la microscopia elettronica.
Microscopia elettronica (EM)
Un altro modo per rilevare i virus è la microscopia elettronica. La disponibilità di questo metodo calando: molti laboratori non la usano più, perché hanno a disposizione strumenti vecchi e il costo e la manutenzione dei nuovi è notevole. Alcuni laboratori dipendono dalle strutture universitarie che forniscono ancora questo servizio, ma con lunghi tempi di diagnosi e costi per il cliente elevati. A differenza della coltura, i microbi non devono restare vitali per la microscopia elettronica. Come la coltura, questo metodo non è specifico (può rilevare molti diversi virus), viene solitamente usata per trovare virus associati con forme enteriche, ed è utile in questa situazione perché questi virus sono di solito eliminati in notevoli quantità (106 particelle virali per rilevarli con EM) e molti di loro sono difficili da coltivare in vitro. In ogni caso, ormai le metodiche molecolari hanno sostituito la microscopia elettronica per individuare i virus.
Rilevamento degli antigeni
Il rilevamento degli antigeni consiste nel trovare le proteine del patogeno. Tale metodo è solitamente specifico, perché si usano gli anticorpi, specialmente i monoclonali, che si legano a una capsula specifica. Se si usano invece i policlonali, questi possono reagire con patogeni diversi. Il beneficio dei policlonali è che più difficilmente si perdono i patogeni, se, ad esempio, c’è stato un cambiamento dell’epitopo che copre anticorpi monoclonali. Il patogeno non deve essere vitale per questa metodica. Metodi di rilevamento dell’antigene includono test con anticorpi fluorescenti, immunoistochimica ed ELISA. In genere, ELISA è più frequentemente usata per il rilevamento degli anticorpi piuttosto che degli antigeni, ma un antigene ben noto ELISA è il BVDV ACE ( cattura antigenica ELISA) per trovare le PI animali. I pro e contro di qualsiasi rilevamento antigenico non vengono qui elencati.
Rilevamento genetico
La reazione della catena della polimerasi (PCR) richiede vari cicli di amplificazione di una regione target del genoma del patogeno. I primer (piccoli pezzi di DNA complementare) sono disegnati per legarsi al patogeno che interessa (e teoricamente a nessun altro). Quindi questa metodica trova solamente ciò che state specificamente cercando nel campione (metodo specifico). I primer sono anche necessari perché avvenga la reazione, poiché la polimerasi del DNA richiede che il primer venga “impressionato” e inizi la sintesi del DNA. La polimerasi del DNA produce solamente parti di DNA complementare, cosi, se un patogeno è a RNA (ad esempio virus a RNA), il materiale genetico deve essere convertito in DNA prima di poter effettuare la PCR; questa operazione viene detta RT (reverse transcription, cioè trascrizione inversa). I nucleotidi liberi (A, T, C, G) vengono inclusi nel mix e aggiunti al DNA del patogeno. I cicli di reazione prevedono la separazione del DNA a doppia elica in singola (denaturazione), il successivo legame dei primer al DNA complementare del patogeno, se presente nel campione (accoppiamento), e l’estensione dei primer da parte della DNA polimerasi (allungamento). Ciascuno di questi passaggi richiede una temperatura differente, il che costituisce il motivo per cui la reazione viene effettuata nel termociclo (cicli con tempi e temperature diverse). L’amplificazione di una porzione del genoma del patogeno, con ciascun ciclo di PCR (ogni ciclo produce una quota doppia), è il risultato. Poiché questa metodica è molto sensibile, è importante che il laboratorio prenda le necessarie precauzioni e usi controlli adeguati, per prevenire o rilevare contaminazioni tra i campioni, onde evitare risultati falsi positivi.
PCR convenzionale
La PCR convenzionale ha diversi nomi: PCR tradizionale, PCR su base gel e PCR finale. Sostanzialmente questo metodo viene usato per visualizzare il prodotto della PCR alla fine del ciclo. Dai diversi cicli di amplificazione deriva una grande serie di prodotti evidenziati nel gel. In base a dove ci si aspetta che i primer leghino il genoma del patogeno, sappiamo quanto prodotto dovrebbe derivarne. Includiamo una scala di DNA nel gel, per determinare la dimensione della banda del prodotto della PCR. Occasionalmente avviene qualcosa di inatteso, amplifichiamo, ad esempio, un prodotto non specifico (quindi non il patogeno che cercavamo), ma che ha le stesse dimensioni di quanto ci si aspettava. Quindi, idealmente, useremo una metodica aggiuntiva (come la sequenziazione) per confermare che prodotto è realmente quello che ci aspettiamo. La sequenziazione viene analizzata in seguito.
PCR real time
Questa è una variazione della PCR convenzionale. Invece di usare un gel per rilevare il prodotto della PCR alla fine della reazione, incorporiamo una sonda fluorescente nella reazione per trovarlo. Questa sonda, se intatta, non produce fluorescenza: la sonda intatta ha una marca fluorescente in un terminale e un relativo blocco nell’altro terminale, che annulla la fluorescenza. Come i primer, la sonda DNA è complementare al DNA del patogeno ed è progettata per legarsi al suo DNA. L’aggiunta di una sonda alla reazione PCR aumenta la specificità del test, perché ora ci sono tre pezzi di DNA complementare che si devono legare al patogeno, anziché solo due. Quando la polimerasi si estende dai primer, prende con sé anche la sonda e la legge nella sequenza. Ciò risulta in una fluorescenza (perché la marca fluorescente viene ora separata dal terminale bloccante), che viene rilevata dal sistema. Pertanto è possibile rilevare l’amplificazione in tempo reale, non appena si forma il prodotto della PCR. C’è comunque sempre una quota di sonde che non restano intatte, e ciò crea alcune fluorescenze di fondo. Il sistema sottrae automaticamente, durante la lettura, tale quota dell’impostazione di base e viene determinata una soglia limite per ogni campione, ovvero il punto in cui viene considerato positivo. Se ci sono molti patogeni nel campione, ci saranno molte sonde spezzate dalla polimerasi, il che si traduce in un aumento della fluorescenza, rispetto a quando ci sono poco patogeni nel campione, ed è anche più veloce (pochi patogeni richiederanno più tempo per produrre la fluorescenza necessaria a raggiungere la soglia). Ovviamente, se il patogeno non è nel campione, la sonda non verrà spezzata, e vedremo solamente una fluorescenza di fondo (generalmente appare come una linea piatta). Il ciclo PCR in cui la fluorescenza passa la soglia, viene detto ciclo soglia (Ct). Un valore inferiore di Ct rappresenta più patogeni nel campione; i valori Ct oltre 35 sono generalmente considerati dubbi/sospetti (e potrebbero non essere riproducibili, perché rappresentano solamente una piccola quota di DNA patogeno nel campione). L’importanza di mettere un valore Ct elevato dipende da ciò che si richiede al test. Ad esempio, se il test viene utilizzato per la sorveglianza in un gruppo clinicamente sano di animali, un valore Ct superiore o uguale a 35 potrebbe essere significativo. Il rilevamento di patogeni non significa necessariamente che essi siano la causa della malattia: potrebbero essere dei secondari, o un isolamento incidentale. In genere, comunque, un Ct basso è un risultato significativo. La PCR real time può essere usata per determinare copie di un campione del patogeno, ma occorre poi fare una curva standard, che solitamente non viene eseguita in diagnostica animale.
Sequenziamento
Esistono due metodiche principali per il sequenziamento: Sanger e Next Generation. Sanger è disponibile da più tempo e viene usata per sequenziare i prodotti PCR che solitamente derivano da PCR convenzionale. Questo metodo richiede anche i primer ed è assai utile se si sa cosa si sta cercando nel campione. Viene comunemente usato per sequenziare geni o porzioni di patogeni ed è è il metodo usato più spesso in diagnostica veterinaria. La sequenziazione può essere usata per batteri speciali difficili da identificare con le prove biochimiche (o Maldi Tof). Può essere usato anche per differenziare infezioni da vaccini vivi modificati o per malattie derivanti da vaccini vettori, differenziandoli da ceppi selvaggi di campo. Una volta che si è ottenuta la sequenza, determiniamo, usando un database statale di sequenze note (Blast), le sequenze ottenute. Sequenziare è anche utile per tracciare dati di epidemiologia molecolare; per fare questo usiamo anche analisi filogenetiche che paragonano sequenze multiple.
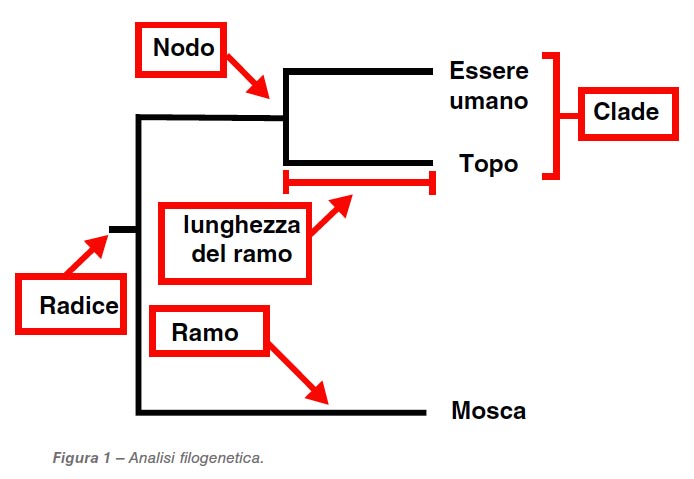
Analisi filogenetica
Gli alberi filogenetici vengono usati per valutare come le sequenze siano correlate (correlazione tra diversi organismi). Visto che sequenziare è diventata una pratica frequente nei laboratori diagnostici, la necessità di paragonare le sequenze e prepararsi a interpretare gli alberi filogenetici sta aumentando. È importante comprendere che non esiste un solo metodo o albero corretto. I metodi usati forniscono anche interferenze; alcuni vengono considerati più accurati di altri, ma diverse metodiche sono comunque accettabili scientificamente. La chiave è arrivare a produrre un albero con radici comuni, che si chiamano cladi (vedi Figura 1), che siano riproducibili. I cladi sono gruppi di organismi che includono un antenato singolo e tutti i discendenti (per i nostri scopi, si tratta delle sequenze correlate). La riproducibilità dei cladi può essere determinata usando il metodo trappola bootstrap, che stima la probabilità di dividere un albero e la relativa correttezza (ovvero che membri di quel clade siano sempre parte dello stesso). La percentuale (ovvero il valore bootstrap) di probabilità che sia vicino al ramo ad esso connesso è importante: maggiore è il numero di nodi (che rappresentano gli avi comuni del clade), meglio è (idealmente se superiore al 75% e oltre) . Occorre un albero in linea con i dati e che si avvicini il più possibile a quanto si è visto in passato, se siamo interessati a conoscere l’evoluzione di un virus o di un batterio. Un’altra cosa da valutare, in un albero filogenetico, è la lunghezza dei rami. Essa rappresenta la quota di variazioni tra un antenato e i suoi discendenti (o, nel nostro caso, la quota di variazioni nucleotidiche): maggiore è la lunghezza, maggiore è il numero di variazioni nucleotidiche. Una chiave è fornita dal numero di variazioni nucleotidiche per sito. La maggioranza di questi varia in modo inapparente (senza variazioni nel rapporto amminoacidi/proteine); così la lunghezza di un ramo non rappresenta per forza variazioni antigeniche, ma solo delle variazioni nucleotidiche. Quindi, solo perché si rilevano variazioni che risultano in un nuovo clade nucleotidico, non significa che i vaccini usati non siano proteggenti. Le variazioni antigeniche sono valutate tramite strumenti differenti.
 Sequenziazioni di nuova generazione (NGS)
Sequenziazioni di nuova generazione (NGS)
Si tratta della nuovissima tecnica molecolare che rivoluziona la diagnostica medica. Questo metodo di sequenziazione varia dal Sanger, perché non richiede primer per le reazioni di sequenziamento e profili di DNA sequenziati in parallelo, nemmeno uno solo per singola reazione. Così, teoricamente, sarebbe possibile sequenziare tutti gli acidi nucleici di un dato campione secondo la cosiddetta metagenomica shotgun o profonda. NGS è più spesso usata per sequenziare genomi di batteri o virus isolati da campioni clinici, o per sequenziare genomi umani, animali o vegetali (WGS). WGS è una tecnica usata dalla FDA e dal Dipartimento di Stato per valutare l’epidemiologia genetica di casi di patogeni di origine animale negli alimenti. La valutazione dei genomi umani sta portando a una medicina di precisione, che si adatta alla composizione genetica del singolo individuo. Nel caso del rilevamento dei patogeni, la metagenomica è molto utile per rilevare organismi; la complicazione sta nel determinare se questi nuovi organismi siano patogeni e responsabili di forme cliniche di campo. Inoltre, visto che non occorrono primer per la NGS, non occorre avere test specifici per tutti i microrganismi presenti sulle liste disponibili. Comunque, per fare un vero test di metagenomica, bisogna spendere diverse centinaia di euro. La quota di DNA o RNA patogeno in un campione (in particolare da un campione di tessuto con diverse cellule animali) è piccola rispetto alla quota di acido nucleico animale: sono quindi necessari milioni di letture di sequenziazioni per trovare il patogeno. Un approccio per ridurre i costi con NGS sta nell’incorporare i primer PCR nel flusso di ciclo: ciò limita la metodica al rilevamento dei soli patogeni per i quali è stato incluso il primer, ma, significativamente, riduce anche il numero di letture necessarie a rilevare il patogeno, e quindi riduce anche i costi. Questo approccio consente comunque ancora il rilevamento di un vasto numero di patogeni in un singolo campione, rendendola così una tecnica assai utile per test multipli. I test multipli PCR vengono frequentemente usati in campioni di tessuti con patogeni, ma sono limitati a pochi target. Inoltre, a differenza di NGS, con la sola PCR non si ottiene alcun dato di sequenziamento, che va fatto tramite una reazione di sequenziamento. NGS è utile perché aree che normalmente sarebbero state target di sequenziamento Sanger, possono ora essere incorporate nel test. In tal modo se si esegue un solo test, rilevare e caratterizzare il tutto viene fatto in una sola volta. Quindi NGS target equivale a PCR multiple con sequenziazione su steroidi. NGS target possono essere eseguite in soli due giorni una volta ricevute dal laboratorio. Mentre lavoravo ad UGA, abbiamo sviluppato un pannello di rilevamento dei patogeni nelle vacche e nei piccoli ruminanti; speriamo di incorporare questo tipo di test nella pratica quotidiana di laboratorio e di sviluppare in futuro ulteriori pannelli.
Bibliografia disponibile su richiesta
Dagli Atti della Midwest Poultry Federeation Convention 2019





